
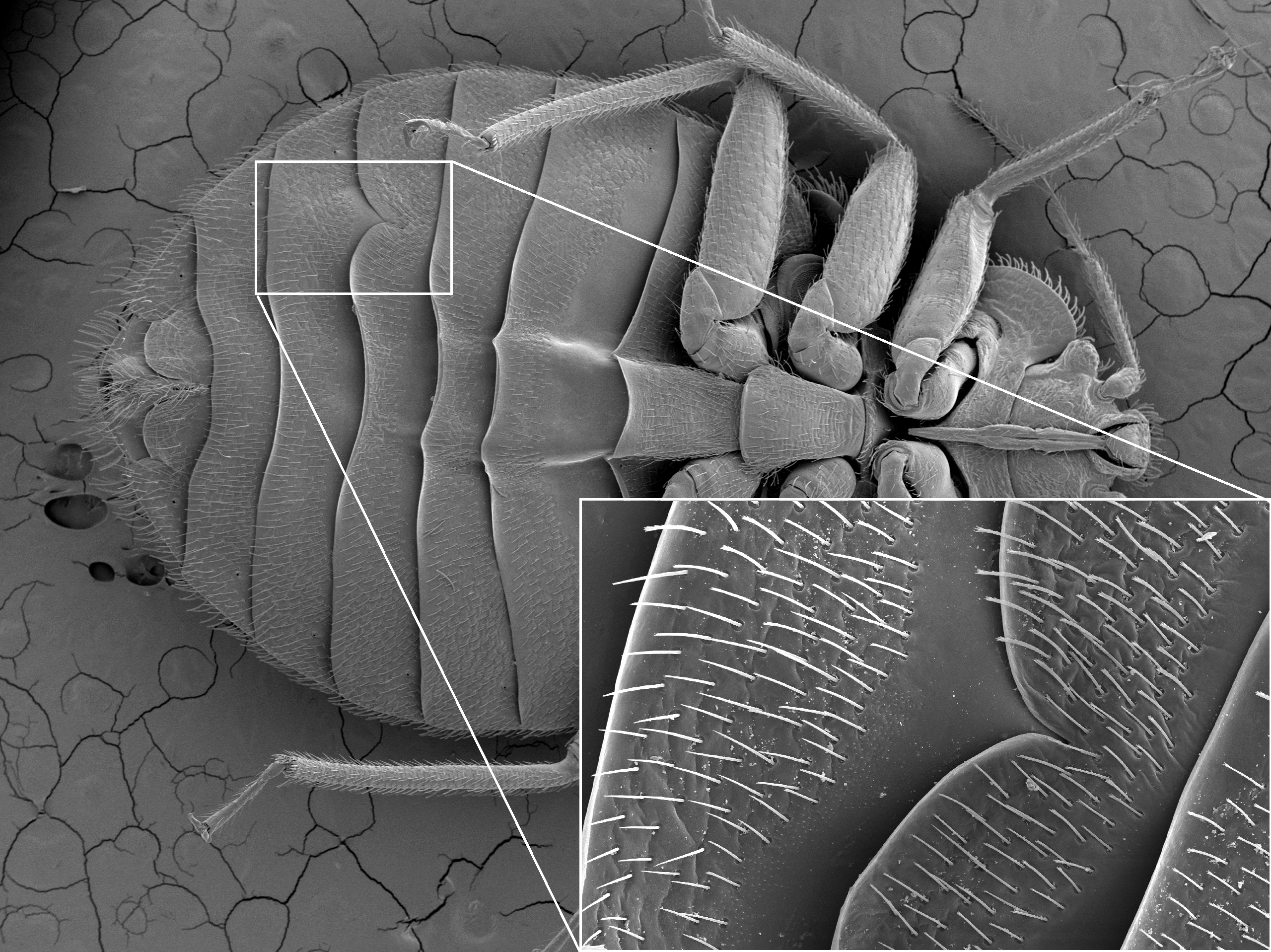
Nejznámějším druhem štěnice je nepochybně pro svůj vztah s člověkem štěnice domácí (Cimex lectularius). V rámci čeledi štěnicovitých je unikátní a rozhodně si zaslouží zvýšenou pozornost i z mnoha dalších důvodů. Má však více než 100 popsaných příbuzných. Všechny druhy jsou obligátní parazité teplokrevných, většinou sociálně žijících obratlovců (netopýři, vlaštovky, rorýsi, člověk). Sají krev hostitelů, v jejichž úkrytech se zdržují. Na tělo hostitele vylézají, jen když se chtějí nasát. Je ovšem pravděpodobné, že s hostiteli příležitostně, ale záměrně cestují (viz náš článek v časopisu Vespertilio)1. Jejich způsob života tak vytváří unikátní strukturu tzv. metapopulací v jednotlivých úkrytech hostitele (blíže v naši publikaci)2. Tato neobvyklá populační struktura pak určuje mechanismy vzniku a fixace adaptací na hostitele, jež jsou zásadním objektem zájmu naší skupiny.
 Cimex lectularius - samice Štěnice domácí je druh původně svázaný s netopýry3. Ve střední Evropě je běžná v synantropních koloniích netopýra velkého. Původní populace v jeskyních možná přežívají na Středním Východě4. Na člověka se specializovala hluboko v pravěku, dávno před vznikem prvních měst (datování na základě mitochondriální DNA – viz náš článek)5. Je doložená spíše z prostředí starověkých civilizací, než ze středověké Evropy. Např. ve Velké Británii se objevila až v 16. století, a to pouze v přístavních městech6. Stejně tak ve Skandinávii či Rusku nebyla prakticky známa ještě v 19. století. S průmyslovou revolucí, urbanizací a především v souvislosti s 1. světovou válkou se však stala pravidelnou součástí domácností. Po druhé světové válce štěnice z většiny zemí prakticky zmizela, především díky používání DDT a pyretroidů. Od konce 90. let se však začíná vracet. Na vině je jistě resistence k insekticidům, možná získaná v důsledku dávného nadužívání DDT. Genetické analýzy však ukazují, že za invazí štěnice nestojí pouze pár kmenů, které díky resistenci přežily2,5,7. Zdá se, že štěnice povstaly spíše z lokálních refugií v důsledku změn chování člověka. Lidé v poslední době mnohem více cestují, navíc zapomněli, jak štěnice vypadá8. Štěnici tak poskytují mnoho příležitostí se šířit. Na konkrétní změny chování člověka, ale i na šíření konkrétních resistentních kmenů, se zaměřuje náš současný populačně genetický projekt.
Cimex lectularius - samice Štěnice domácí je druh původně svázaný s netopýry3. Ve střední Evropě je běžná v synantropních koloniích netopýra velkého. Původní populace v jeskyních možná přežívají na Středním Východě4. Na člověka se specializovala hluboko v pravěku, dávno před vznikem prvních měst (datování na základě mitochondriální DNA – viz náš článek)5. Je doložená spíše z prostředí starověkých civilizací, než ze středověké Evropy. Např. ve Velké Británii se objevila až v 16. století, a to pouze v přístavních městech6. Stejně tak ve Skandinávii či Rusku nebyla prakticky známa ještě v 19. století. S průmyslovou revolucí, urbanizací a především v souvislosti s 1. světovou válkou se však stala pravidelnou součástí domácností. Po druhé světové válce štěnice z většiny zemí prakticky zmizela, především díky používání DDT a pyretroidů. Od konce 90. let se však začíná vracet. Na vině je jistě resistence k insekticidům, možná získaná v důsledku dávného nadužívání DDT. Genetické analýzy však ukazují, že za invazí štěnice nestojí pouze pár kmenů, které díky resistenci přežily2,5,7. Zdá se, že štěnice povstaly spíše z lokálních refugií v důsledku změn chování člověka. Lidé v poslední době mnohem více cestují, navíc zapomněli, jak štěnice vypadá8. Štěnici tak poskytují mnoho příležitostí se šířit. Na konkrétní změny chování člověka, ale i na šíření konkrétních resistentních kmenů, se zaměřuje náš současný populačně genetický projekt.
 Cimex hemipterus - samiceŠtěnice tropická (Cimex hemipterus) měla a má podobný osud jako štěnice domácí. V Evropě je nacházena spíše příležitostně, je vázána na teplé oblasti. Původní populace na netopýrech jsou známy z Indie9. Z většiny území zmizela také díky DDT. To, že tento druh v posledních letech znovu obsadil stejná území6, také dokládá, že na vině návratu štěnic jsou globální změny chování lidské společnosti, nikoliv pouze šíření resistentních kmenů.
Cimex hemipterus - samiceŠtěnice tropická (Cimex hemipterus) měla a má podobný osud jako štěnice domácí. V Evropě je nacházena spíše příležitostně, je vázána na teplé oblasti. Původní populace na netopýrech jsou známy z Indie9. Z většiny území zmizela také díky DDT. To, že tento druh v posledních letech znovu obsadil stejná území6, také dokládá, že na vině návratu štěnic jsou globální změny chování lidské společnosti, nikoliv pouze šíření resistentních kmenů.
 Cimex pipistrelli - samecŠtěnice netopýří (Cimex pipistrelli) sdílí ve středoevropském prostoru se štěnicí domácí kolonie netopýra velkého. Je však běžná u mnoha dalších netopýrů, především u netopýra rezavého10. Tento druh má ve zvyku obsazovat panelové domy. Díky tomu se se štěnicí netopýří občas potká i člověk (blíže náš článek)11. Trvalé populace na člověku však nejsou známy. Z naší zkušenosti vyplývá, že na lidské krvi obtížně dokončuje svůj vývojový cyklus.
Cimex pipistrelli - samecŠtěnice netopýří (Cimex pipistrelli) sdílí ve středoevropském prostoru se štěnicí domácí kolonie netopýra velkého. Je však běžná u mnoha dalších netopýrů, především u netopýra rezavého10. Tento druh má ve zvyku obsazovat panelové domy. Díky tomu se se štěnicí netopýří občas potká i člověk (blíže náš článek)11. Trvalé populace na člověku však nejsou známy. Z naší zkušenosti vyplývá, že na lidské krvi obtížně dokončuje svůj vývojový cyklus.
 Cimex hirundinis - samice (vlevo) a samecŠtěnice ptačí (Cimex hirundinis) byla pro svůj vzhled dříve řazena do samostatného rodu Oeciacus. Naše genetické analýzy12 však ukázaly, že se vyvinula v rámci rodu Cimex, tedy skupiny štěnic svázaných s netopýry. Svůj vzhled změnila díky vztahu s ptáky. V Evropě je běžná v hnízdech jiřiček, v menší míře vlaštovek a rorýsů. Často se tak dostane do lidských obydlí. Na člověku se však zřejmě nedokáže ani nasát. Dlouhodobé asociace jsou tedy vyloučené.
Cimex hirundinis - samice (vlevo) a samecŠtěnice ptačí (Cimex hirundinis) byla pro svůj vzhled dříve řazena do samostatného rodu Oeciacus. Naše genetické analýzy12 však ukázaly, že se vyvinula v rámci rodu Cimex, tedy skupiny štěnic svázaných s netopýry. Svůj vzhled změnila díky vztahu s ptáky. V Evropě je běžná v hnízdech jiřiček, v menší míře vlaštovek a rorýsů. Často se tak dostane do lidských obydlí. Na člověku se však zřejmě nedokáže ani nasát. Dlouhodobé asociace jsou tedy vyloučené.
Ostatní druhy štěnic tvoří více či méně lokální populace téměř na všech kontinentech. Trvalejší asociace s člověkem zřejmě vytváří africký Leptocimex boueti9. Před druhou světovou válkou byla v Novém Světě běžným škůdcem na drůbeži štěnice Haematosiphon inodorus. Nyní ji lze najít pouze u jejího původního hostitele – dravých ptáků. Je však možné, že je, ač historicky, čtvrtým druhem štěnice spjatým s lidmi, konkrétně s indiány kmene Hopi. Pro tuto myšlenku existuje celkem přesvědčivá lingvistická podpora13.
Ektoparazitický způsob života je pouze jeden z unikátních znaků štěnic. Další nápadnou vlastností je jejich způsob páření – traumatická inseminace. Samec samici páří pomocí modifikovaného přívěsku, nikoliv penisu, do membrány mezi články zadečku. Zde má samice také specializovaný orgán, který se vyvinul jako adaptace na mechanické zranění i možné infekce. I přesto je však kopulace pro samici stresující. Mezidruhová kopulace může být až fatální, jak bylo dokumentováno při páření štěnice domácí se štěnicí tropickou14,15. Na efekt mezidruhového páření se zaměřujeme ve vztahu štěnice domácí a štěnice netopýří. Tyto dva druhy si konkurují v prostředí kolonií netopýra velkého a traumatická inseminace by mohla být jedním z mechanismů mezidruhového boje.
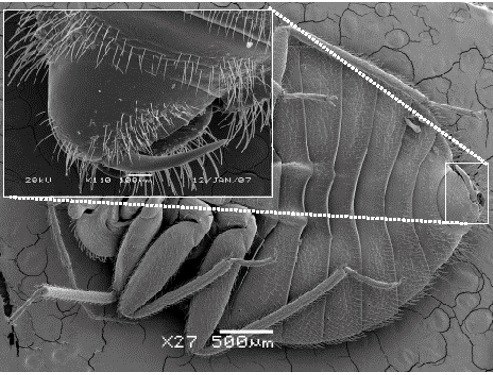 Samec - paramera Samec - paramera |
 Samice - paragenitální orgán Samice - paragenitální orgán |
Štěnice jsou také neobvyklé v dědičnosti genetického materiálu. Variabilní počet pohlavních chromosomů je u štěnice domácí a několika dalších druhů známý již dlouho. Teprve nedávno naši kolegové z PřF UK prokázali, že tyto nadpočetné chromosomy nemusí vznikat fragmentací16, 17. Jaderný genom štěnice má tedy proměnlivou velikost.
 Larva štěniceŠtěnice, především štěnice domácí, jsou ale zcela unikátní v dědičnosti mitochondriálního genomu (mtDNA). Biparentální dědičnost mtDNA je napříč živočišnou říší dokumentována často. S výjimkou jedné skupiny škeblí jde ale ve všech případech o mizivé poměry otcovské mtDNA vůči mateřské. Většinou k únikům otcovské mtDNA dochází při hybridizaci příbuzných druhů, nebo vzdálených populací jednoho druhu. Tyto případy naplňují příslovečné výjimky, potvrzující univerzální pravidlo dědičnosti mtDNA po matce. Štěnice však zřejmě dědí mitochondrie od obou rodičů velmi často (blíže v našem článku)18 a představují tak teprve druhý takový případ mezi živočichy. Podobně jako u škeblí, varianty mtDNA štěnic spolu rekombinují, jak ukazuje bádání našeho kolegy Warrena Bootha (nepublikováno).
Larva štěniceŠtěnice, především štěnice domácí, jsou ale zcela unikátní v dědičnosti mitochondriálního genomu (mtDNA). Biparentální dědičnost mtDNA je napříč živočišnou říší dokumentována často. S výjimkou jedné skupiny škeblí jde ale ve všech případech o mizivé poměry otcovské mtDNA vůči mateřské. Většinou k únikům otcovské mtDNA dochází při hybridizaci příbuzných druhů, nebo vzdálených populací jednoho druhu. Tyto případy naplňují příslovečné výjimky, potvrzující univerzální pravidlo dědičnosti mtDNA po matce. Štěnice však zřejmě dědí mitochondrie od obou rodičů velmi často (blíže v našem článku)18 a představují tak teprve druhý takový případ mezi živočichy. Podobně jako u škeblí, varianty mtDNA štěnic spolu rekombinují, jak ukazuje bádání našeho kolegy Warrena Bootha (nepublikováno).
1/ Balvín, O. et al. Transport of bugs of the genus Cimex (Heteroptera: Cimicidae) by bats in western Palaearctic. Vespertilio 16, 43 –54 (2012).
2/ Booth, W., Balvín, O., Vargo, E. L., Vilímová, J. & Schal, C. Host association drives genetic divergence in the bed bug, Cimex lectularius. Mol. Ecol. 24, 980–992 (2015).
3/ Horváth, G. La distribution géographique des cimicides et l´origine des punaises des lits. in Extrait du IXe Congres International de Zoologie Tenu a Monaco 294–299 (1913).
4/ Povolný, D. & Usinger, R. L. The discovery of a possibly aboriginal population of the bed bug (Cimex lectularius Linnaeus, 1958). Acta Musei Morav. Sci. Nat. 51, 237–242 (1966).
5/ Balvín, O., Munclinger, P., Kratochvíl, L. & Vilímova, J. Mitochondrial DNA and morphology show independent evolutionary histories of bedbug Cimex lectularius (Heteroptera: Cimicidae) on bats and humans. Parasitol. Res. 111, 457,–469 (2012).
6/ Naylor, R., Balvín, O., Delaunay, P. & Akhoundi, M. The bed bug resurgence in Europe and Russia. in Advances in the Biology and Management of Modern Bed Bugs (John Wiley and Sons, 2018).
7/ Szalanski, A. L., Austin, J. W., McKern, J. A., Steelman, C. D. & Gold, R. E. Mitochondrial and Ribosomal Internal Transcribed Spacer 1 Diversity of Cimex lectularius (Hemiptera: Cimicidae). J. Med. Entomol. 45, 229–236 (2008).
8/ Reinhardt, K., Harder, A., Holland, S., Hooper, J. & Leake-Lyall, C. Who knows the bed bug? Knowledge of adult bed bug appearance increases with people age in three counties of Great Britain. J. Med. Entomol. 45, 956–958 (2008).
9/ Usinger, R. L. Monograph of Cimicidae. (Entomological Society of America, 1966).
10/ Balvín, O., Bartonička, T., Simov, N., Paunovic, M. & Vilímová, J. Distribution and host relations of species of the genus Cimex on bats in Europe. Folia Zool. 63, (2014).
11/ Balvín, O. & Bartonička, T. Cimicids and bat hosts in the Czech and Slovak Republics: ecology and distribution. Vespertilio 17, 23–36 (2014).
12/ Balvín, O., Roth, S. & Vilímová, J. Molecular evidence places the swallow bug genus Oeciacus Stal within the bat and bed bug genus Cimex Linnaeus (Heteroptera: Cimicidae): The genus Oeciacus within the genus Cimex. Syst. Entomol. 40, 652–665 (2015).
13/ Reinhardt, K. Which bed bug the Hopi know? (A present for Robert Leslie Usinger’s 100th birthday). Am. Entomol. 58, 58c–59 (2012).
14/ Omori, N. Experimental studies on the cohabitation and crossing of bed-bugs (Cimex lectularius L. and C. hemipterus F.). Preliminary report. in VII. International Kongres der Entomologie (ed. Uschmann, G.) 2, 895–915 (1939).
15/ Walpole, D. E. Cross-mating studies between two species of bedbugs (Hemiptera: Cimicidae) with a description of a marker of interspecific mating. South Afr. J. Sci. 84, 215–216 (1988).
16/ Sadílek, D., Urfus, T. & Vilímova, J. Am I really so smashed? in Zoologické dny (2018).
17/ Sadílek, D., Urfus, T., Vilímová, J., Hadrava, J., Suda, J. (2019) Nuclear Genome Size in Contrast to Sex ChromosomeNumber Variability in the Human Bed Bug,Cimex lectularius (Heteroptera: Cimicidae). Cytometry Part A.
18/ Robinson, G., Balvin, O., Schal, C., Vargo, E. & Booth, W. Extensive mitochondrial heteroplasmy in natural populations of a resurging human pest, the bed bug. J. Med. Entomol. accepted, (2015).